
Отбор группы особей обладающих полезными признаками

Выделяют три формы естественного отбора: движущий, стабилизирующий и разрывающий (дизруптивный).
Движущий отбор — естественный отбор, который действует при направленном изменении условий внешней среды.
Особи с признаками, которые соответствуют изменяющейся среде, получают преимущества. При этом другие проявления признаков не сохраняются. В результате в популяции из поколения к поколению происходит сдвиг среднего значения признака в определённом направлении. Формируется новая средняя норма вместо существующей, переставшей соответствовать новым условиям.
Схема действия движущего отбора
В результате движущего отбора одни признаки исчезают, а другие формируются. Если отбор действует в одном направлении длительное время, то он может привести к превращению популяции в новый вид.
Примеры:
- изменение окраски у берёзовой пяденицы в промышленных районах (промышленный меланизм);
- редукция глаз у крота, пищеварительной системы у паразитических ленточных червей;
- увеличение длины корня у склерофитов;
- превращение передних конечностей в роющие у разных групп животных при освоении почвы как среды обитания;
- формирование у бактерий устойчивости к антибиотикам;
- снижение чувствительности паразитических грибов к ядохимикатам;
- изменение строения конечностей у предков лошади в связи с освоением открытых пространств.
Изменение конечностей у предков лошади
Стабилизирующий отбор — естественный отбор, действие которого направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака.
Действует в постоянных условиях среды.
Направлен на поддержание ранее сложившегося среднего признака или свойства. Сохраняет приспособленность вида, устраняя резкие отклонения выраженности признака от средней нормы, тем самым предохраняет сложившийся генотип от разрушающего действия мутационного процесса.
Схема действия стабилизирующего отбора
Отбор в пользу средних значений был обнаружен по множеству признаков.
Примеры:
- гибель во время сильной бури птиц, имеющих слишком короткие и слишком длинные крылья;
- более частая гибель при рождении или в первые недели жизни новорожденных млекопитающих с очень низким и очень высоким весом;
- сохранение в популяциях зайцев особей с оптимальной длиной конечностей;
- сохранение у растений, опыляющихся насекомыми, определённого строения цветка, соответствующего размерам насекомых;
- существование реликтовых видов организмов, сохранившихся в неизменном виде на протяжении миллионов лет (кистепёрая рыба латимерия, гинкго, гаттерия и др.).
Кистепёрая рыба латимерия
Дизруптивный (разрывающий) отбор — естественный отбор, при котором сохраняются крайние варианты признака, а убираются его средние значения.
Схема действия дизруптивного отбора
В результате может появиться несколько новых форм из одной исходной. Дизруптивный отбор способствует возникновению и поддержанию разнообразия популяций. Он приводит к дивергенции (расхождению признаков) и образованию нескольких видов из одного исходного.
Примеры:
- формирование на сенокосных лугах двух рас погремка — раннецветущей и позднецветущей;
- существование весенних и осенних форм и видов грибов из-за повторяющейся в середине лета засухи;
- возникновение разных подвидов и видов синиц в связи с пищевой специализацией;
- возникновение разных видов клевера.
Клевер средний
Клевер луговой
Источник
Селекция (лат. selectio – выбирать) – наука и отрасль практической деятельности, направленная на создание новых сортов растений,
пород животных и штаммов микроорганизмов, обладающих полезными для человека свойствами.
Этими полезными свойствами могут быть размер и форма плодов, урожайность, удойность у коров, устойчивость к факторам внешней
среды (к засушливому климату, к морозу).

Основы селекции
В основе селекции лежит способность генотипа живых организмов к изменениям, что происходит главным образом за счет комбинативной и
мутационной изменчивости. В процессе селекции происходит искусственный отбор организмов с полезными для человека свойствами и их размножение.
В результате множества последовательных скрещиваний, в конце концов, селекционерам удается достичь желаемой цели: вывести гибридов
с нужными признаками.
Мутационная изменчивость существует благодаря мутациям – случайным ненаправленным изменениям генотипа. Благодаря мутациям, к примеру, возник безалкалоидный сорт люпина. И.В. Мичуриным на яблоне сорта Антоновка Могилевская были обнаружены необычайно крупные плоды,
ветвь с которым послужила для появления нового сорта – Антоновки шестистограммовой. Эти плоды – результат произошедшей в
естественных условиях мутации соматических клеток.

“Сколько ждать этой естественной мутации?” – спросите вы. Может один день, а может и 100, и 10000 лет – всем властвует случайность.
На наш век может не выпасть удача, а мы такого допустить не можем! 🙂
Именно по этой причине в селекции растений часто используются искусственно вызванные мутации – авто- и аллополиплоидию.
Автополиплоидия
Автополиплоидия – кратное (4n,6n,8n) увеличение исходного набора хромосом, который характерен для
особей вида.
Автополиплоидия возникает в результате обработки почек колхицином, который нарушает образование нитей веретена деления, и, соответственно, нарушает расхождение хромосом при митозе, в результате чего набор хромосом в клетке оказывается удвоенным. Таким способом получают полиплоиды – сорта растений, обладающие повышенной
урожайностью.
Существуют различные тетраплоидные сорта свеклы, мака, кукурузы и других сельскохозяйственных культур, которые отличаются большими размерами плодов.

Аллополиплоидия
Аллополиплоидия (греч. állos — другой и polýploos — многократный) – соединение в клетках организма хромосомного набора
от разных видов или родов, в результате которого образуется гибридная зигота.
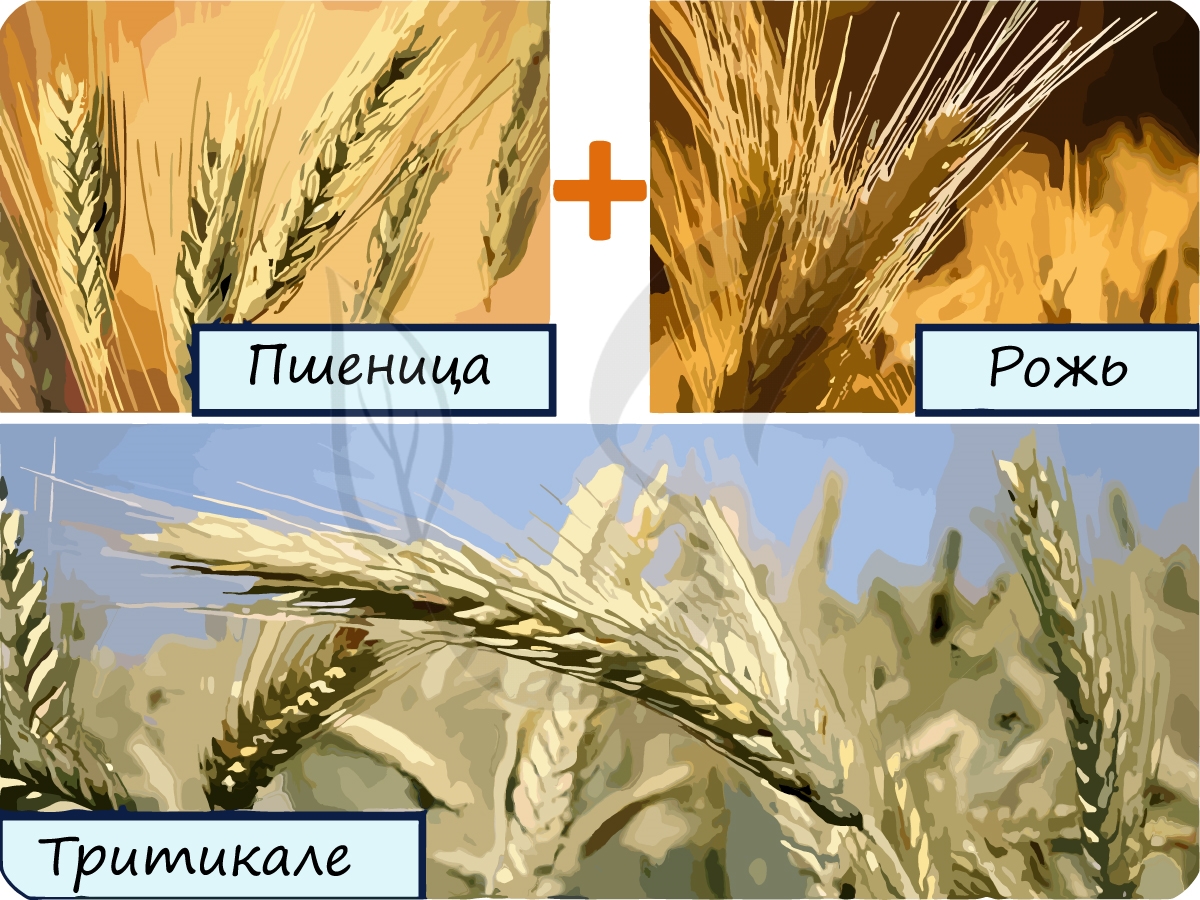
Благодаря аллополиплоидии получают новые сорта растений. Наиболее известным примером является гибрид ржи и пшеницы – тритикале. Некоторые межвидовые гибриды табака обладают повышенной устойчивостью к возбудителям заболеваний мучнистой росы, табачной мозаики.
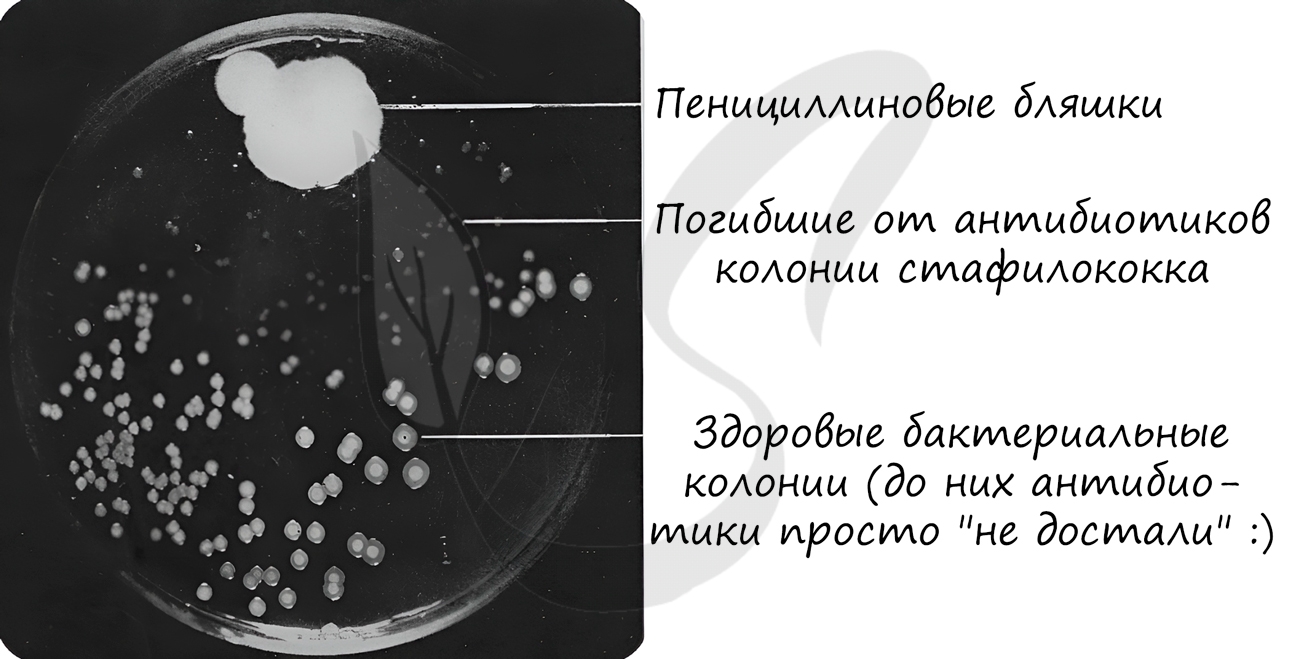
В рамках биотехнологии разработаны методы, с помощью которых стало возможным создание бактерий, синтезирующих полезные для
человека белки, многие из которых используются как лекарства: аминокислоты, антибиотики, инсулин.
Скрещивание особей в селекции
Каждое скрещивание как сдача новых карт: может повезет, а может и нет. Вполне возможно, что особь унаследует полезные признаки от родителей и сможет передать их своим потомкам, всегда есть и шанс того, что появятся новые полезные для человека признаки, равно как и шанс, что ничего полезного из проводимого скрещивания не выйдет.
Возможны несколько вариантов скрещивания:
- Близкородственное скрещивание (инбридинг – от англ. in — внутри + breeding — разведение)
- Неродственное скрещивание (аутбридинг – от англ. out — вне + breeding — разведение)
- Отдаленная гибридизация
Близкородственное скрещивание в течение нескольких поколений приводит к переходу генов в гомозиготное состояние, вследствие чего
потомство ослабевает и становится более подвержено наследственным заболеваниям.
Замечу, что под инбридингом подразумевают близкородственное скрещивание животных. Для самоопыления у растений существует иной
термин – инцухт.
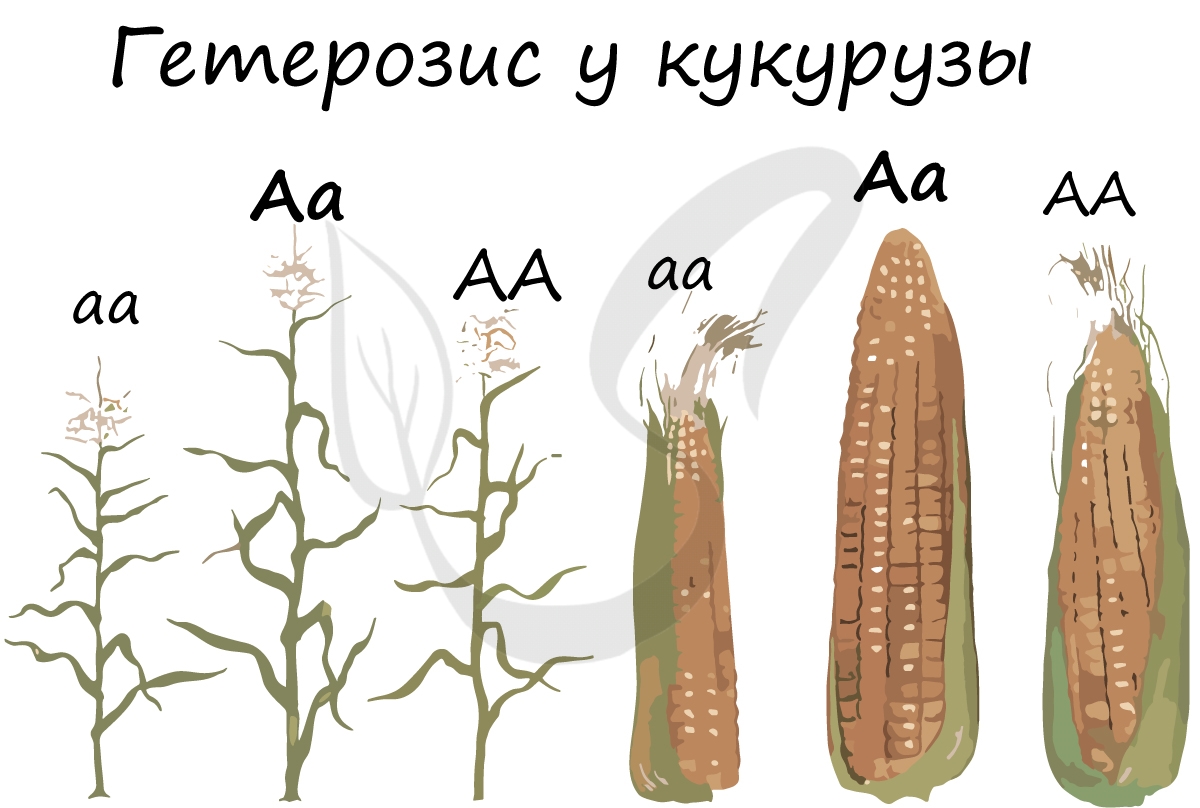
В селекции инбридинг применяют для выведения чистых линий (гомозиготных особей – aa, AA, bb, BB), которые используются, например, для
анализирующего скрещивания. Инбридинг использовался при выведении абсолютно всех пород животных, и в настоящее время активно используется
в питомниках для выведения нужных пород животных (кошек, собак и т.д.)

Аутбридинг заключается в скрещивании неродственных особей, которые могут принадлежать к одному сорту, породе, виду или роду. Аутбридинг ведет к явлению гетерозиса – получения гетерозисных форм, которые превосходят родительских
особей по ряду признаков.
Гетерозис – явление увеличения жизнеспособности особей у гибридов, которые получены при скрещивании двух чистых линий. Такой эффект
связан с переходом генов в гетерозиготное состояние, что повышает выживаемость организмов, плодовитость, и множество других полезных свойств.
Применение отдаленной гибридизации заключается в скрещивании особей, принадлежащих к разным родам и видам. Такие особи обладают крайне
полезными для человека свойствами, но часто бесплодны (стерильны).
Известным примером отдаленной гибридизации является мул – гибрид осла (самца) и лошади (самки). Отличаются большой выносливостью и
работоспособностью, живут до 40 лет, обладают хорошим иммунитетом к заболеваниям, не требовательны в корме и уходе.
Обратный пример: гибрид ослицы (самки) и жеребца (самца) – лошак. Встречаются гораздо реже по сравнению с мулом, так как обладают меньшей
выносливостью и работоспособностью. В большинстве случаев бесплодны.
Отбор в селекции
Отбор в селекции осуществляет человек с единственной целью: размножить особей с нужными и полезными признаками, свойствами. Очевидно, что такой
отбор называется искусственным, в противовес естественному отбору, главный критерий которого – приспособленность.
Отбор может осуществляться двумя способами:
- Массовый отбор
- Индивидуальный отбор
Отбор организмов исключительно на основе внешних данных (фенотипа). Основным критерием для человека служит проявление признака:
размер плодов, цвет лепестков, цвет листьев и т.д. Этот вид отбора характеризуется массовостью и быстротой.
В результате массового отбора формируется группа особей, которые обладают нужными и полезными для человека признаками. В дальнейшем
они подвергаются размножению.

Выборочный отбор и сохранение особей с ценными для человека признаками. В ходе индивидуального отбора оценивается не только фенотип,
но и генотип, вследствие чего данный вид отбора занимает большее время, но оказывается более эффективен.
Индивидуальный отбор требует оценки потомства от выбранной особи в ряду поколений. Иногда подобный отбор применяют у самоопыляемых
растений: пшеницы, ячменя – с целью получения чистых линий. Как было сказано ранее, чистые линии характеризуются гомозиготностью и являются исходным материалом для селекции.

© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Есте́ственный отбо́р — основной фактор эволюции, в результате действия которого в популяции увеличивается число особей, обладающих более высокой приспособленностью (наиболее благоприятными признаками), в то время как количество особей с неблагоприятными признаками уменьшается. В свете современной синтетической теории эволюции естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов. Естественный отбор — единственная известная причина адаптаций, но не единственная причина эволюции. К числу неадаптивных причин относятся генетический дрейф, поток генов и мутации.
Термин «Естественный отбор» популяризовал Чарльз Дарвин, сравнивая данный процесс с искусственным отбором, современной формой которого является селекция. Идея сравнения искусственного и естественного отбора состоит в том, что в природе так же происходит отбор наиболее «удачных», «лучших» организмов, но в роли «оценщика» полезности свойств в данном случае выступает не человек, а среда обитания. К тому же, материалом как для естественного, так и для искусственного отбора являются небольшие наследственные изменения, которые накапливаются из поколения в поколение[1].
Механизм естественного отбора[править | править код]
В процессе естественного отбора закрепляются мутации, увеличивающие приспособленность организмов к окружающей их среде. Естественный отбор часто называют «самоочевидным» механизмом, поскольку он следует из таких простых фактов, как:
- Организмы производят потомков больше, чем может выжить;
- В популяции этих организмов существует наследственная изменчивость;
- Организмы, имеющие разные генетические черты, имеют различную выживаемость и способность размножаться.
Такие условия создают конкуренцию между организмами в выживании и размножении и являются минимально необходимыми условиями для эволюции посредством естественного отбора[2]. Таким образом, организмы с наследственными чертами, которые дают им конкурентное преимущество, имеют большую вероятность передать их своим потомкам, чем организмы с наследственными чертами, не имеющими подобного преимущества[3].
Центральное понятие концепции естественного отбора — приспособленность организмов[4]. Приспособленность определяется как способность организма к выживанию и размножению в существующей окружающей среде. Это определяет размер его генетического вклада в следующее поколение[4]. Однако главным в определении приспособленности является не общее число потомков, а число потомков с данным генотипом (относительная приспособленность)[5]. Например, если потомки успешного и быстро размножающегося организма слабые и плохо размножаются, то генетический вклад и, соответственно, приспособленность этого организма будут низкими[4].
Если какая-либо аллель увеличивает приспособленность организма больше, чем другие аллели этого гена, то с каждым поколением доля этой аллели в популяции будет расти. То есть, отбор происходит в пользу этой аллели. И наоборот, для менее выгодных или вредных аллелей — их доля в популяциях будет снижаться, то есть отбор будет действовать против этих аллелей[6]. Важно отметить, что влияние определённых аллелей на приспособленность организма не является постоянным — при изменении условий окружающей среды вредные или нейтральные аллели могут стать полезными, а полезные вредными[7].
Естественный отбор для черт, которые могут изменяться в некотором диапазоне значений (например, размер организма), можно разделить на три типа[8]:
- Движущий (направленный) отбор — изменения среднего значения признака в течение долгого времени, например увеличение размеров тела;
- Дизруптивный отбор — отбор на крайние значения признака и против средних значений, например, большие и маленькие размеры тела;
- Стабилизирующий отбор — отбор против крайних значений признака, что приводит к уменьшению дисперсии признака.
Частным случаем естественного отбора является половой отбор, субстратом которого является любой признак, который увеличивает успешность спаривания за счёт увеличения привлекательности особи для потенциальных партнёров[9]. Черты, которые эволюционировали за счёт полового отбора, особенно хорошо заметны у самцов некоторых видов животных. Такие признаки, как крупные рога, яркая окраска, с одной стороны могут привлекать хищников и понижать выживаемость самцов[10], а с другой это уравновешивается репродуктивным успехом самцов с подобными ярко выраженными признаками[11].
Отбор может действовать на различных уровнях организации — таких, как гены, клетки, отдельные организмы, группы организмов и виды[12][13]. Причём отбор может одновременно действовать на разных уровнях[14]. Отбор на уровнях выше индивидуального, например, групповой отбор, может приводить к кооперации (см. Эволюция#Кооперация)[15].
Формы естественного отбора[править | править код]
Существуют разные классификации форм отбора. Широко используется классификация, основанная на характере влияния форм отбора на изменчивость признака в популяции.
Движущий отбор[править | править код]
Движущий отбор — форма естественного отбора, которая действует при направленном изменении условий внешней среды. Описали Дарвин и Уоллес. В этом случае особи с признаками, которые отклоняются в определённую сторону от среднего значения, получают преимущества. При этом иные вариации признака (его отклонения в противоположную сторону от среднего значения) подвергаются отрицательному отбору. В результате в популяции из поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию).
Две разноокрашенные бабочки берёзовой пяденицы на одном дереве
Примером действия движущего отбора является «индустриальный меланизм» у насекомых. «Индустриальный меланизм» представляет собой резкое повышение доли меланистических (имеющих тёмную окраску) особей в тех популяциях насекомых (например, бабочек), которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц, а тёмные — хуже. В XX веке в ряде районов доля тёмноокрашенных бабочек в некоторых хорошо изученных популяциях берёзовой пяденицы в Англии достигла 95 %, в то время как впервые тёмная бабочка (morfa carbonaria) была отловлена в 1848 году.
Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определённом направлении, перемещая соответственно и норму реакции. Например, при освоении почвы как среды обитания у различных неродственных групп животных конечности превратились в роющие.
Стабилизирующий отбор[править | править код]
Стабилизирующий отбор — форма естественного отбора, при которой его действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака. Понятие стабилизирующего отбора ввёл в науку и проанализировал И. И. Шмальгаузен.
Описано множество примеров действия стабилизирующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детёнышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорождённые с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорождённые со средним весом. Учёт размера крыльев у воробьёв, погибших после бури в 50-х годах под Ленинградом, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
Дизруптивный отбор[править | править код]
Дизруптивный (разрывающий) отбор — форма естественного отбора, при которой условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дарвин описывал действие дизруптивного отбора, считая, что он лежит в основе дивергенции, хотя и не мог привести доказательств его существования в природе. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования.
Одна из возможных в природе ситуаций, в которой вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам.
Примером дизруптивного отбора является образование двух рас у погремка большого на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая.
Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись только особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора.
Половой отбор[править | править код]
Половой отбор — это естественный отбор на успех в размножении. Выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важным компонентом является привлекательность для особей противоположного пола. Дарвин назвал это явление половым отбором. «Эта форма отбора определяется не борьбой за существование в отношениях органических существ между собой или со внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола». Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения, значительно выше, чем их недостатки для выживания.
Распространены две гипотезы о механизмах полового отбора.
- Согласно гипотезе «хороших генов», самка «рассуждает» следующим образом: «Если данный самец, несмотря на яркое оперение и длинный хвост, сумел не погибнуть в лапах хищника и дожить до половой зрелости, значит он обладает хорошими генами, которые позволили ему это сделать. Следовательно, его стоит выбрать в качестве отца своих детей: он передаст им свои хорошие гены». Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков.
- Согласно гипотезе «привлекательных сыновей», логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более усиливается. Процесс идёт по нарастающей, пока не достигнет предела жизнеспособности.
При выборе самцов самки не задумываются о причинах своего поведения. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме — оно идёт на водопой, потому что чувствует жажду. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами — им нравятся яркие хвосты. Те, кому инстинкт подсказывал иное поведение, не оставили потомства. Логика борьбы за существование и естественного отбора — логика слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы.
Роль естественного отбора в эволюции[править | править код]
Чарльз Дарвин полагал естественный отбор основной движущей силой эволюции, в современной синтетической теории эволюции он также является основным регулятором развития и адаптации популяций, механизмом возникновения видов и надвидовых таксонов, хотя накопление в конце XIX — начале XX века сведений по генетике, в частности обнаружение дискретного характера наследования фенотипических признаков, привело к тому[источник не указан 3016 дней], что некоторые исследователи стали отрицать важность естественного отбора, и в качестве альтернативы предлагали концепции, базирующиеся на оценке фактора мутации генотипа как чрезвычайно важного. Авторы таких теорий постулировали не постепенный, а очень быстрый (в течение нескольких поколений) скачкообразный характер эволюции (мутационизм Гуго де Фриза, сальтационизм Рихарда Гольдшмитда и другие, менее известные концепции). Открытие известных корреляций среди признаков родственных видов (закон гомологических рядов) Н. И. Вавилова подтолкнуло некоторых исследователей к формулировке очередных «антидарвиновских» гипотез об эволюции — таких, как номогенез, батмогенез, автогенез, онтрогенез и прочие.
В 1920—1940-е годы в эволюционной биологии, у тех, кто отвергнул идею Дарвина об эволюции путём естественного отбора, (иногда теории, в которых уделялось большое значение естественному отбору называли «селекционистскими») возродился интерес к этой теории благодаря пересмотру классического дарвинизма в свете относительно молодой науки генетики. Разработанная в результате этого синтетическая теория эволюции, часто некорректно называемая неодарвинизмом, помимо прочего, опирается на количественный анализ частоты аллелей в популяциях, изменяющейся под влиянием естественного отбора. Ведутся споры[источник не указан 3016 дней], где люди с радикальным подходом в качестве аргумента против синтетической теории эволюции, а также роли естественного отбора утверждают, что «открытия последних десятилетий в различных областях научного знания — от молекулярной биологии с её теорией нейтральных мутаций Мотоо Кимуры и палеонтологии с её теорией прерывистого равновесия Стивена Джея Гоулда и Найлза Элдриджа (в которой вид понимается как относительно статическая фаза эволюционного процесса) до математики с её теорией бифуркаций и фазовых переходов — свидетельствуют о недостаточности классической синтетической теории эволюции для адекватного описания всех аспектов биологической эволюции»[источник не указан 3016 дней]. Дискуссия о роли различных факторов в эволюции началась более 30 лет назад и продолжается по сегодня, и иногда высказывается, что «эволюционная биология (подразумевая под этим теорию эволюции, разумеется) подошла к необходимости своего очередного, третьего синтеза»[источник не указан 3016 дней].
Интересные факты[править | править код]
Дарвин долго не решался опубликовать свою работу, так как наблюдал «проблему муравьёв»[17].
На примере рабочего муравья мы имеем насекомое, чрезвычайно отличающееся от своих родителей, тем не менее, абсолютно бесплодное и, следовательно, не могущее передавать из поколения в поколение приобретённые модификации строения или инстинктов. Можно задать хороший вопрос — насколько возможно согласовать этот случай с теорией естественного отбора?
— Происхождение видов (1859)
Дарвин предполагал, что отбор может применяться не только к индивидуальному организму, но и к семейству. Он также говорил, что, возможно, в той или иной мере это может объяснять и поведение людей. Он оказался прав, однако предоставить более расширенное представление этой концепции стало возможно только после появления генетики. Первый набросок «теории родственного отбора» сделал английский биолог Уильям Гамильтон в 1963 году, который первым предложил рассматривать естественный отбор не только на уровне индивида или целого семейства, но и на уровне гена[18].
См. также[править | править код]
- Искусственный отбор
- Селекция
- Эволюционизм
- Давление отбора
Примечания[править | править код]
- ↑ Естественный отбор // Казахстан. Национальная энциклопедия. — Алматы: Қазақ энциклопедиясы, 2005. — Т. II. — ISBN 9965-9746-3-2.
- ↑ Солдбриг, 1982, с. 43—47.
- ↑ Futuyma, 2005, p. 251—252.
- ↑ 1 2 3 Orr H. A. Fitness and its role in evolutionary genetics // Nature Reviews Genetics. — 2009. — Vol. 10, no. 8. — P. 531—539. — doi:10.1038/nrg2603. — PMID 19546856.
- ↑ Haldane J. B. S. The theory of natural selection today (англ.) // Nature. — 1959. — Vol. 183, no. 4663. — P. 710—713. — PMID 13644170.
- ↑ Lande R., Arnold S. J. The measurement of selection on correlated characters (англ.) // Evolution. — Wiley-VCH, 1983. — Vol. 37, no. 6. — P. 1210—1226. — doi:10.2307/2408842.
- ↑ Futuyma, 2005.
- ↑ Грант, 1991, Глава 14.
- ↑ Andersson M., Simmons L. W. Sexual selection and mate choice (англ.) // Trends in Ecology & Evolution. — Cell Press, 2001. — Vol. 21, no. 6. — P. 296—302. — doi:10.1016/j.tree.2006.03.015. — PMID 16769428.
- ↑ Kokko H., Brooks R., McNamara J. M., Houston A. I. The sexual selection continuum // Proceedings of the Royal Society. Biological Sciences. — 2002. — Vol. 269, no. 1498. — P. 1331—1340. — doi:10.1098/rspb.2002.2020. — PMID 12079655.
- ↑ Hunt J., Brooks R., Jennions M. D., Smith M. J., Bentsen C. L., Bussière L. F. High-quality male field crickets invest heavily in sexual display but die young (англ.) // Nature. — 2004. — Vol. 432, no. 7020. — P. 1024—1027. — doi:10.1038/nature03084. — PMID 15616562.
- ↑ Okasha S. . Evolution and the Levels of Selection (англ.). — Oxford: Oxford University Press, 2007. — 263 p. — ISBN 0-19-926797-9.
- ↑ Mayr E. The objects of selection // Proc. Nat. Acad. Sci. USA. — 1997. — Т. 94, no. 6. — P. 2091—2094. — PMID 9122151.
- ↑ Maynard Smith J. The units of selection // Novartis Foundation Symposium. — 1998. — Vol. 213. — P. 211—217. — PMID 9653725.
- ↑ Gould S. J., Lloyd E. A. Individuality and adaptation across levels of selection: how shall we name and generalise the unit of Darwinism? // Proc. Nat. Acad. Sci. USA. — 1999. — Т. 96, no. 21. — P. 11904–11909. — PMID 10518549.
- ↑ Darwin, Charles (1859). On the Origin of Species (1st edition). Chapter 4, page 88. «And this leads me to say a few words on what I call Sexual Selection. This depends …» https://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F373&pageseq=12
- ↑ Этология. Ру: Моральное животное, 2 часть
- ↑ Родственный отбор. Эволюция кооперации и альтруизма.
Литература[править | править код]
- Грант В. . Эволюционный процесс. Критический обзор эволюционной теории / Под ред. Б. М. Медникова. — М.: Мир, 1991. — 488 с. — ISBN 5-03-001432-2. Архивная копия от 9 апреля 2012 на Wayback Machine
- Дарвин Ч. Глава IV // О происхождении видов в царствах животном и растительном путём естественного подбора родичей или о сохранении усовершенствованных пород в борьбе за существование / пер. С. А. Рачинский — СПб.: Глазунов, Александр Ильич (издатель), 1864. — С. 65—108.
- Солбриг О., Солбриг Д. . Популяционная биология и эволюция. — М.: Мир, 1982. — 488 с.
- Futuyma D. J. Evolution. — Sunderland: Sinauer Associates, 2005. — 543 p. — ISBN 0-878-93187-2.
Ссылки[править | править код]
- «Формы естественного отбора» — статья с хорошо известными примерами: цвет бабочек, устойчивость людей к малярии и прочее
- On the Origin of Species by Charles Darwin — Chapter 4, Natural Selection
- Natural Selection — Modeling for Understanding in Science Education, University of Wisconsin
- Natural Selection from University of Berkeley education website
- T. Ryan Gregory: Understanding Natural Selection: Essential Concepts and Common Misconceptions (недоступная ссылка) Evolution: Education and Outreach
Источник